Dzielić:



Warstwy kory mózgowej, komórki i funkcje (z obrazami)
The kora mózgowa lub kora mózgowa jest tkanką nerwową pokrywającą powierzchnię półkul mózgowych. Mówiąc o innej formie, stanowi najbardziej nadrzędny obszar mózgu.
Ta struktura mózgu osiąga maksymalny rozwój u naczelnych, jest mniej rozwinięta u innych zwierząt i jest związana z rozwojem działań poznawczych i intelektualnych bardziej złożonych.
Kora mózgowa jest podstawowym obszarem mózgu dla funkcjonowania ludzi. W tym regionie wykonywane są takie funkcje, jak percepcja, wyobraźnia, myśl, osąd lub decyzja.
Anatomicznie składa się z szeregu cienkich warstw składających się z istoty szarej, które znajdują się nad szerokim zbiorem ścieżek istoty białej.
Kora mózgowa przybiera zwiniętą formę, więc gdyby miała się rozszerzyć, miałaby bardzo rozległą masę. W szczególności badania sugerują, że całkowita powierzchnia kory mózgowej może składać się z około 2500 centymetrów kwadratowych.
Podobnie, ta duża masa mózgu charakteryzuje się tym, że zawiera ogromną liczbę neuronów w swoim wnętrzu. Ogólnie rzecz biorąc, postuluje się, że w korze mózgowej jest około 10 miliardów neuronów, które wytwarzałyby około 50 bilionów synaps.
Główne cechy kory mózgowej wyjaśniono poniżej. Określono jego warstwy, neurony i organizację funkcjonalną, a funkcje, które są wykonywane w tym obszarze mózgu, są weryfikowane.
Indeks
- 1 Charakterystyka kory mózgowej
- 2 warstwy
- 2.1 Warstwa molekularna
- 2.2 Zewnętrzna warstwa ziarnista
- 2.3 Zewnętrzna warstwa piramidalna
- 2.4 Wewnętrzna warstwa ziarnista
- 2.5 Warstwa węzłów chłonnych
- 2.6 Wielowarstwowa warstwa
- 3 Organizacja funkcjonalna
- 3.1 Obszary wrażliwe
- 3.2 Obszary motorowe
- 3.3 Obszary stowarzyszenia
- 4 komórki nerwowe
- 4.1 Komórki piramidalne
- 4.2 Komórki gwiaździste
- 4.3 Komórki wrzeciona
- 4.4 Cajal poziome komórki
- 4.5 komórki Martinottiego
- 5 referencji
Charakterystyka kory mózgowej
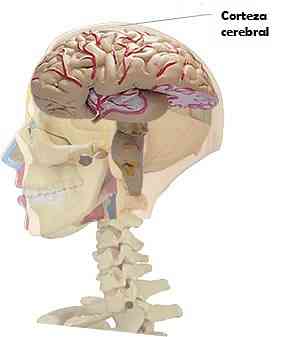
Kora mózgowa zwierząt ssaków jest reprezentowana przez warstwę istoty szarej, która pokrywa dwie półkule mózgowe.
Składa się z bardzo złożonej struktury, w której różne organy czuciowe są reprezentowane w określonych obszarach lub obszarach, które nazywane są pierwotnymi obszarami sensorycznymi.
Każdy z pięciu zmysłów posiadanych przez człowieka (wzrok, dotyk, zapach, smak i dotyk) rozwija się w określonym obszarze kory. Oznacza to, że każda modalność zmysłowa ma wyznaczony obszar w korze mózgowej.
Oprócz obszarów czuciowych kora mózgowa ma również wiele wtórnych obszarów somatycznych, asocjacyjnych i motorycznych. W tych obszarach rozwijają się układy aferentne korowe i asocjacyjne, co powoduje uczenie się, pamięć i zachowanie.
W tym sensie, kora mózgowa jest uważana za szczególnie istotny region przy rozwijaniu najwyższej aktywności ludzkiego mózgu.
Najbardziej zaawansowane i skomplikowane procesy ludzi, takie jak wnioskowanie, planowanie, organizacja lub stowarzyszenie, są przeprowadzane w różnych obszarach kory mózgowej.
Z tego powodu kora mózgowa stanowi strukturę, która uzyskuje maksymalną złożoność z ludzkiej perspektywy. Kora mózgowa jest wynikiem powolnego procesu ewolucyjnego, który mógł rozpocząć się ponad 150 milionów lat temu.
Warstwy
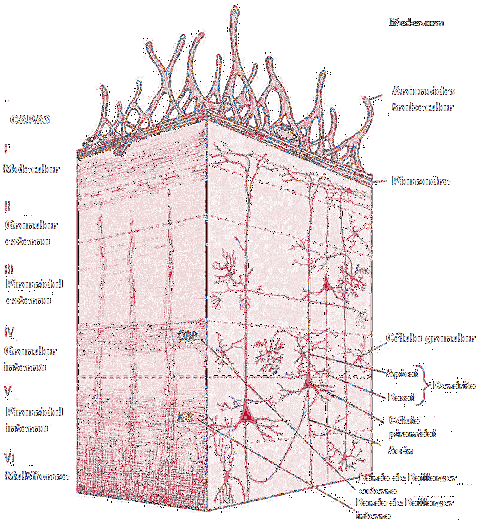
Główną cechą kory mózgowej jest to, że składa się z różnych warstw istoty szarej. Warstwy te tworzą strukturę skorupy i definiują jej strukturę strukturalną i funkcjonalną.
Ponadto warstwy kory mózgowej charakteryzują się nie tylko definiowaniem z strukturalnego punktu widzenia, ale także z perspektywy filogenetycznej.
Oznacza to, że każda z warstw kory mózgowej odpowiada innej chwili ewolucyjnej. Na początku gatunku ludzkiego mózg był mniej rozwinięty, a kora miała mniej warstw.
Poprzez ewolucję gatunku warstwy te wzrastają, co wiąże się ze wzrostem zdolności poznawczych i intelektualnych ludzi w miarę upływu czasu.
Warstwa molekularna
Warstwa molekularna, znana również jako warstwa pleksi, jest najbardziej powierzchownym obszarem kory mózgowej, a zatem najnowszym początkiem.
Składa się z gęstej sieci włókien nerwowych zorientowanych stycznie. Włókna te pochodzą z dendrytów komórek piramidalnych i wrzecionowatych, aksonów komórek gwiaździstych i Martinottiego.
Włókna doprowadzające, które pochodzą ze wzgórza, asocjacji i spoidła, można również znaleźć w warstwie molekularnej. Będąc najbardziej powierzchownym obszarem kory, w warstwie molekularnej powstaje duża liczba synaps między różnymi neuronami.
Zewnętrzna warstwa ziarnista
Zewnętrzna warstwa ziarnista jest drugim najbardziej powierzchownym obszarem kory i leży poniżej warstwy molekularnej. Zawiera dużą liczbę małych komórek piramidalnych i gwiaździstych.
Dendryty komórek zewnętrznej warstwy ziarnistej trafiają do warstwy molekularnej, a aksony wchodzą do głębszych warstw kory mózgowej. Z tego powodu zewnętrzna warstwa ziarnista jest połączona z różnymi obszarami kory.
Zewnętrzna warstwa piramidalna
Zewnętrzna warstwa piramidalna, jak sugeruje jej nazwa, składa się z komórek piramidalnych. Charakteryzuje się nieregularnym kształtem, tzn. Rozmiar warstwy wzrasta od granicy powierzchni do najgłębszego ograniczenia.
Dendryty neuronów warstwy piramidalnej przechodzą do warstwy molekularnej, a aksony przemieszczają się jako projekcja, asocjacja lub włókna spoidłowe do istoty białej znajdującej się między warstwami kory mózgowej.
Wewnętrzna warstwa ziarnista
Wewnętrzna warstwa ziarnista składa się z komórek gwiaździstych, które są ułożone w bardzo zwartej formie. Ma wysokie stężenie poziomo ułożonych włókien zwanych zewnętrznym pasmem Baillargera.
Warstwa zwężająca
Warstwa zwojowa lub wewnętrzna warstwa piramidalna zawiera bardzo duże i średniej wielkości komórki piramidalne. Podobnie zawierają dużą liczbę włókien ułożonych poziomo, które tworzą wewnętrzne pasmo Baillarger.
Warstwa wielopostaciowa
Wreszcie wielowarstwowa warstwa, znana również jako warstwa komórek polimorficznych, zasadniczo zawiera komórki wrzecionowate. Podobnie zawiera zmodyfikowane komórki piramidalne zawierające trójkątne lub jajowate ciało komórki.
Wiele włókien nerwowych wielowarstwowej warstwy wchodzi w leżącą pod nią istotę białą i łączy warstwę z obszarami pośrednimi.
Organizacja funkcjonalna
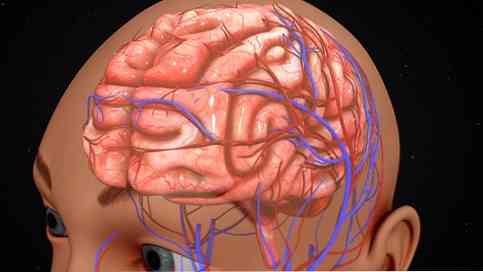
Kora mózgowa może być również zorganizowana zgodnie z działaniami prowadzonymi w każdym regionie. W tym sensie pewne obszary kory mózgowej przetwarzają specyficzne sygnały o charakterze wrażliwym, motorycznym i asocjacyjnym..
Obszary wrażliwe
Obszary sensoryczne są obszarami kory mózgowej, które otrzymują informacje o wrażliwym charakterze i są ściśle związane z percepcją.
Informacja uzyskuje dostęp do kory mózgowej głównie przez tylną połowę obu półkul mózgowych. Obszary pierwotne zawierają najbardziej bezpośrednie połączenia z obwodowymi receptorami czuciowymi.
Z drugiej strony, drugorzędne obszary sensoryczne i asocjacyjne zwykle sąsiadują z obszarami pierwotnymi. Ogólnie rzecz biorąc, otrzymują informacje zarówno z głównych obszarów skojarzenia, jak i z niższych obszarów mózgu..
Głównym zadaniem obszarów stowarzyszenia i obszarów drugorzędnych jest integracja wrażliwych doświadczeń w celu wygenerowania wzorców rozpoznawania i zachowania. Główne wrażliwe obszary kory mózgowej to:
- Główny obszar somatosensoryczny (obszary 1, 2 i 3).
- Główny obszar wizualny (obszar 17).
- Główny obszar słuchowy (obszar 41 i 42).
- Główny obszar smaku (obszar 43).
- Pierwotny obszar węchowy (obszar 28).
Obszary motoryczne
Obszary motoryczne znajdują się w przedniej części półkul. Są odpowiedzialni za inicjowanie procesów mózgowych związanych z ruchem i powodujących takie działania.
Najważniejsze obszary motoryczne to:
- Główny obszar silnika (obszar 4).
- Obszar języka ćwiczeń (obszar 44 i 45).
Obszary stowarzyszenia
Obszary asocjacji kory mózgowej korelują z bardziej złożonymi funkcjami integracyjnymi. Regiony te wykonują czynności takie jak procesy pamięci i poznania, zarządzanie emocjami oraz rozwój rozumowania, woli lub osądu.
Podobnie obszary stowarzyszenia odgrywają szczególnie ważną rolę w rozwoju osobowości i cech charakteru ludzi. Podobnie jest to obszar mózgu niezbędny do określenia inteligencji.
Obszary stowarzyszenia obejmują pewne obszary ruchowe, jak również określone regiony sensoryczne.
Komórki nerwowe

Kora mózgowa ma w sobie wiele różnych komórek. W szczególności w tym obszarze mózgu określono pięć różnych typów neuronów.
Komórki piramidalne
Komórki piramidalne to neurony charakteryzujące się kształtem piramidy. Większość tych komórek ma średnicę od 10 do 50 mikrometrów.
Istnieją jednak również duże komórki piramidalne. Są one znane jako komórki Betza i mogą mieć średnicę do 120 mikrometrów.
Zarówno małe komórki piramidalne, jak i duże komórki piramidalne znajdują się w motorycznej wstępnej cyrkulacji i głównie wykonują czynności związane z ruchem.
Komórki gwiaździste
Komórki gwiaździste, znane również jako komórki ziarniste, są małymi neuronami. Zwykle mają średnicę około 8 mikrometrów i mają kształt wielokąta.
Komórki wrzecionowe
Komórki wrzecionowate to neurony, których pionowa oś wzdłużna znajduje się na powierzchni. Są one skoncentrowane głównie w głębszych warstwach korowych mózgu.
Akson tych neuronów pochodzi z dolnej części ciała komórki i jest skierowany w stronę istoty białej jako projekcja, asocjacja lub włókno spoidłowe..
Cajal poziome komórki
Poziome komórki cajal to małe wrzecionowate komórki zorientowane poziomo. Znajdują się w najbardziej powierzchownych warstwach kory mózgowej i odgrywają kluczową rolę w rozwoju tego regionu mózgu.
Ten typ neuronów odkrył i opisał Ramón y Cajal pod koniec XIX wieku, a kolejne badania wykazały, jak istotne komórki mają koordynować aktywność neuronalną.
Aby osiągnąć swoją pozycję w korze mózgowej, poziome komórki cajala muszą migrować w sposób skoordynowany podczas embriogenezy mózgu. Oznacza to, że neurony te przemieszczają się z miejsca urodzenia do powierzchni kory mózgowej.
W odniesieniu do wzoru molekularnego tych neuronów, Victor Borrell i Óscar Marín z Institute of Neuroscience w Alicante, wykazali, że poziome komórki cajala przedstawiają orientację warstw neuronalnych kory podczas rozwoju embrionalnego.
W rzeczywistości dyspersja tych komórek pochodzi z początkowych etapów rozwoju embrionalnego. Komórki rodzą się w różnych regionach mózgu i migrują na powierzchnię mózgu, aby całkowicie ją pokryć.
Wreszcie niedawno wykazano, że błony oponowe pełnią inne funkcje niż te ochronne, które przypuszczano na początku. Opony służą jako podłoże lub ścieżka poziomych komórek cajala dla ich stycznej migracji wzdłuż powierzchni skorupy.
Komórki Martinottiego
Ostatnimi neuronami, które tworzą aktywność neuronalną kory mózgowej, są dobrze znane komórki Martinottiego. Składają się z małych wielopostaciowych neuronów obecnych na wszystkich poziomach kory mózgowej.
Te neurony zawdzięczają swoją nazwę Carlo Martinotti, studenckiemu naukowcowi Camilo Golgiemu, który odkrył istnienie tych komórek kory mózgowej.
Komórki Martinottiego charakteryzują się tym, że są wielobiegunowymi neuronami z krótkimi, drzewiastymi dendrytami. Są one rozsiewane przez kilka warstw kory mózgowej i wysyłają aksony do warstwy molekularnej, gdzie powstają arborizacje aksonalne.
Ostatnie badania tych neuronów wykazały, że komórki Martinottiego uczestniczą w mechanizmie hamowania mózgu.
W szczególności, gdy neuron piramidalny (który jest najczęstszym typem neuronu w korze mózgowej) zaczyna nadmiernie ekscytować, komórki Martinottiego zaczynają przekazywać sygnały hamujące do otaczających komórek nerwowych..
W tym sensie wywnioskowano, że padaczka może być silnie związana z niedoborem komórek Martinottiego lub niedoborem aktywności tych neuronów. W tych chwilach nerwowa transmisja mózgu nie jest już regulowana przez te komórki, co powoduje brak równowagi w funkcjonowaniu kory..
Referencje
- Abeles M, Goldstein MH. Architektura funkcjonalna w pierwotnej korze słuchowej kota. Organizacja i organizacja kolumn według głębokości. J Neurophysiol 1970; 33: 172-87.
- Blasdel GG, Lund JS. Zakończenie aksonów doprowadzających w korze prążkowanej makaka. J Neurosci 1983; 3: 1389-413.
- Chang HT. Neurony korowe ze szczególnym uwzględnieniem dendrytów wierzchołkowych. Cold Spring Harb Symp Quant Biol 1952; 17: 189-202.
- Od Felipe J. Chandelier komórki i padaczka. Brain 1999; 122: 1807-22.
- Ramón y Cajal S. Neue Darstellung vom histologischen Bau des Centralnerevensystem. Arch Anat Physiol 1893: 319-428.
- Rubenstein JLR, Rakic P. Genetyczna kontrola rozwoju korowego. Cereb Cortex 1999; 9: 521-3.